


Foi no passado dia 7 de Outubro de 2019 que ficámos a conhecer os três investigadores galardoados com o tão ambicionado Nobel da Medicina e Fisiologia. William Kaelin, Peter Ratcliffe e Gergg Semenza foram premiados pelas suas descobertas relacionadas com os mecanismos moleculares que permitem às células detectar e adaptar-se aos níveis de oxigénio disponíveis.

William Kaelin é especialista em Medicina Interna e Oncologia, e médico no Hospital de Brigham. Concomitantemente exerce actividade académica na Faculdade de Medicina da Universidade de Harvard, e de investigação no Instituto de Cancro de Dana-Farber.
Peter Ratcliffe especializou-se em Nefrologia e é actualmente professor na Universidade de Oxford, para além de director do departamento de investigação clínica no Instituto Francis Crick, em Londres.
Gregg Semenza, especialista em Pediatria e pós-graduado em Genética Médica, é professor na Universidade Johns Hopkins e director do departamento de investigação vascular para a Engenharia Celular no Instituto com o mesmo nome. O seu laboratório dedica-se sobretudo ao estudo dos mecanismos moleculares por trás da angiogénese (processo de formação de novos vasos sanguíneos) e remodelação vascular na doença cardiovascular isquémica.
O oxigénio, representado pela fórmula química O2, é um gás que constitui cerca de 1/3 da atmosfera terrestre, sendo essencial à vida. Os animais precisam de oxigénio para converter os alimentos e nutrientes em energia. No entanto, os seus níveis podem diminuir substancialmente em todo o organismo – por exemplo, em elevadas altitudes ou durante o exercício físico – ou numa área específica do mesmo – como no local de uma ferida. Este estado de redução dos níveis de oxigénio (conhecido como hipoxia), levam a processos de adaptação nas células, permitindo ao organismo sobreviver mesmo em condições adversas. A maioria destes mecanismos, como a angiogénese, a formação de células sanguíneas, ou a glicólise/fermentação anaeróbica, pretende rentabilizar a utilização do escasso oxigénio existente, bem como produzir energia através de processos poupadores de oxigénio.
Desta forma, ao longo da evolução, os organismos foram-se desenvolvendo por forma a garantir uma suplementação de oxigénio suficiente para todos os tecidos e células. O corpo carotídeo, presente nas carótidas (artérias do pescoço) contém células especializadas que detectam os níveis de oxigénio no sangue e que posteriormente comunicam directamente com o cérebro, controlando, entre outras coisas, o ritmo respiratório e cardíaco.
Para além do mecanismo de rápida adaptação à hipoxia controlado pelo corpo carotídeo, existe também uma resposta desencadeada pelo aumento dos níveis da hormona eritropoietina (EPO). A EPO é uma hormona produzida no rim que aumenta a produção de glóbulos vermelhos (mecanismo conhecido como eritropoiese) na medula óssea, quando os níveis de oxigénio no organismo são baixos. Desta forma, aumentamos os “transportadores” de oxigénio na corrente sanguínea, melhorando o aporte deste gás aos diferentes tecidos. A importância do controlo hormonal da eritropoiese já é conhecida há alguns anos, mas o processo pelo qual este mecanismo era controlado pelos níveis de oxigénio no sangue ainda permanecia um mistério.
Semenza e os seus colaboradores, estudaram o gene EPO e o processo pelo qual este é regulado. Utilizando ratinhos geneticamente modificados, conseguiram demonstrar que existem determinados segmentos de DNA localizados proximamente ao gene EPO, que medeiam a resposta à hipoxia.
Ratcliffe e os seus colaboradores, também estudaram a regulação do gene EPO dependente dos níveis de oxigénio.
Ambas as equipas descobriram que existem mecanismos de detecção de oxigénio presentes em quase todos os tecidos, e não apenas nas células renais onde a EPO é normalmente produzida.
Em 1990, utilizando culturas de células hepáticas, Semenza descobriu um complexo proteico capaz de se ligar a um segmento específico de DNA, dependente dos níveis de oxigénio no sangue. Ele chamou a este complexo Factor Induzido pela Hipoxia (HIF – hipoxia-inducible factor).
Alguns anos mais tarde, em 1995, descobriu também que este complexo HIF é composto por 2 factores de transcrição (proteínas capazes de se ligar ao DNA e que controlam a sua transcrição em RNA), actualmente chamadas HIF-1a e ARNT. Descobriu também que um deles é sensível aos níveis de oxigénio no sangue (HIF-1a), enquanto que o outro não (ARNT). Percebeu assim que o HIF-1a é o componente do HIF que regula a resposta aos níveis de oxigénio.
Ratcliffe, por sua vez, veio provar que os níveis de HIF-1a eram regulados por alterações na estabilidade da proteína, e não por alterações na transcrição genética ou na síntese proteica.
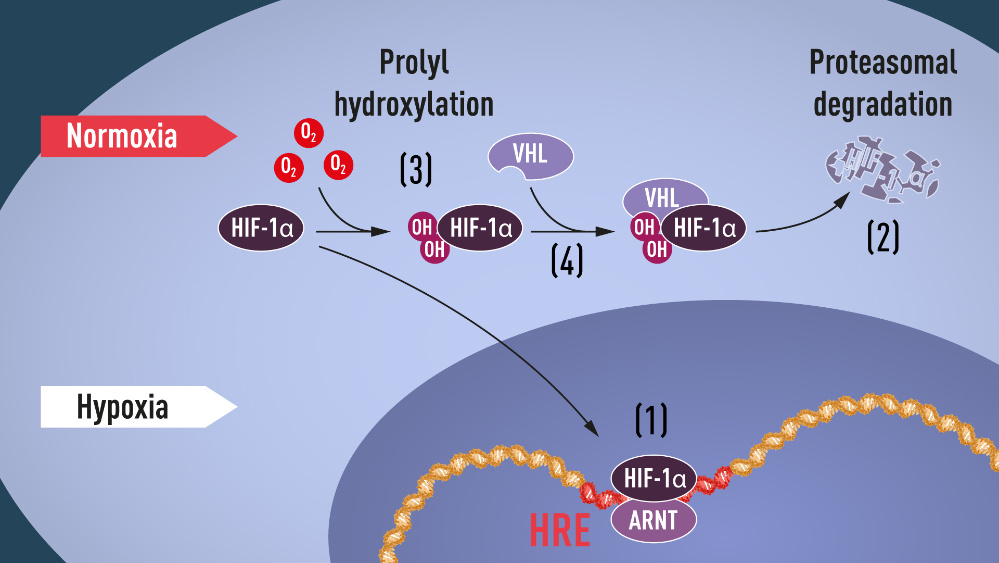
Quando os níveis de oxigénio no sangue estão elevados, um pequeno péptido chamado ubiquitina liga-se ao HIF-1a. A ubiquitina funciona como uma “etiqueta” para proteínas destinadas à degradação. Desta forma, o HIF-1a é destruído por um componente celular chamado proteossoma, e os seus níveis nas células caem. No entanto, quando os níveis de oxigénio baixam, o HIF-1a está de certa forma protegido, e os seus níveis nas células aumentam, permitindo a sua ligação ao gene EPO. A forma como a ubiquitina se ligava ao HIF-1a também permanecia um mistério.
Kaelin estudou durante vários anos uma síndrome hereditária chamada doença de von Hippel-Lindau. Esta doença genética é caracterizada por mutações a nível do gene supressor de tumor VHL, levando ao aumento hereditário do risco de aparecimento de vários cancros.
Em 1995, Kailen publicou pela primeira vez a sequência do gene VHL, e conseguiu provar que este gene codifica uma proteína que previne o aparecimento de cancro.
Kaelin e os seus colaboradores também demonstraram, de forma inesperada, que as células cancerígenas com um gene VHL não funcional expressam de forma anormal níveis elevados de genes reguladores de hipoxia. No entanto, quando o gene VHL é reintroduzido nas células cancerígenas, os níveis normais são restaurados. Este foi um achado importante que veio mostrar que o VHL está de certa forma envolvido no controlo da resposta à hipoxia.
Em 1996, a mesma equipa demonstrou que um determinado número de genes relacionados com o HIF estavam sobrexpressos em conjuntos de células com o VHL mutado.
Este achado sugeriu que a via de resposta ao oxigénio dependente do HIF e a via de tumorogénese relacionada com o VHL, podiam estar ligadas de alguma forma.
Alguns anos mais tarde, em 1999, Ratcliffe e os seus colaboradores demonstraram que o VHL consegue interagir fisicamente com o HIF-1a e que é necessário para a sua degradação na presença de níveis elevados ou normais de oxigénio no sangue. Foram ainda mais longe e demonstraram que o complexo VHL está directamente envolvido na proteólise (processo de degradação de proteínas por enzimas) do HIF-1a, ao agir como um componente de reconhecimento para o complexo de ubiquitina.
Estas descobertas vieram mostrar de forma clara e conclusiva a relação existente entre o HIF-1a e o VHL.
Os mesmos protagonistas, Kaelin e Ratcliffe, seguindo o trilho de investigação em redor do gene VHL e do factor de transcrição HIF-1a, vieram, simultaneamente, em 2001, mostrar que a regulação do HIF-1a pelo VHL depende da sua hidroxilação, uma modificação covalente que é por si só dependente da presença de oxigénio.
As duas equipas mostraram que em níveis elevados ou normais de oxigénio, são adicionados grupos hidroxilo (OH) em 2 posições específicas do HIF-1a e que esta modificação proteica, chamada propil-hidroxilação, aumenta a afinidade do complexo VHL ao HIF-1a. Após a ligação do VHL ao HIF-1a, este é finalmente reconhecido pela ubiquitina que “rotula” o factor de transcrição para degradação. Por outro lado, na ausência de oxigénio, a hidroxilação do HIF-1a não é possível, e o VHL não é capaz de o reconhecer. Consequentemente, o HIF-1a não é ubiquitinado, evitando assim a degradação proteossómica e acumulando-se na célula com activação transcipcional do programa dos genes induzidos por hipoxia.
Todo este mecanismo explica como os níveis de oxigénio controlam de forma rápida a degradação do HIF-1a.
Estas descobertas abriram caminho para novas estratégias promissoras de combate a várias doenças como é o caso de alguns tipos de anemia e do cancro.
Doentes com Insuficiência Renal Crónica sofrem frequentemente de anemia devido à diminuição da expressão de EPO. Ao descobrirmos formas de aumentar a expressão do gene HIF-1a, como por exemplo através da criação de inibidores específicos da hidroxilação deste factor de transcrição, podemos ajudar a tratar estes doentes.
Nos cancros, os mecanismos fisiológicos regulados pelo oxigénio são utilizados para estimular a angiogénese, e para reinventar o metabolismo das células cancerígenas, favorecendo a sua proliferação. Ao baixar a expressão do gene HIF-1a podemos tentar limitar a angiogénese verificada, e reduzir assim a suplementação de nutrientes e outros factores de crescimento às células cancerígenas.
Sem dúvida que todas estas descobertas contribuíram para um avanço imensurável no nosso conhecimento acerca dos mecanismos de adaptação do organismo a diferentes níveis de oxigénio, e que prometem guiar e estimular nos próximos anos a investigação farmacológica direccionada ao tratamento de múltiplas patologias, entre elas a anemia e o cancro.



